Noyau: morphologie, structure, composition chimique, fonctions et signification du noyau
Noyau: morphologie, structure, composition chimique, fonctions et signification du noyau!
La caractéristique la plus importante d'une cellule vue au microscope est le noyau. En 1700, Leeuwenhoek découvrit à l'origine qu'il s'agissait de corps rétractiles au centre des globules sanguins de sang de Saumon.
Ces structures, qui devaient être des noyaux, ont été observées avec les lentilles simples qu’il a trouvées lors de ses loisirs. En 1781, Fontana observa des corps ovales similaires à l'intérieur des cellules cutanées d'une anguille. Le botaniste écossais Robert Brown en donna une description complète en 1835.
C'est une structure presque universelle de cellules à un moment ou à un autre de leur cycle de vie, bien que des cellules telles que les tubes criblés de plantes vasculaires et les globules rouges de mammifères puissent perdre leur noyau au moment où elles se différencient complètement.
Le noyau n'est pas homogène, mais contient des structures filiformes qui ont été découvertes quelques années plus tard. Ces structures pourraient être obtenues avec des colorants à l'aniline, raison pour laquelle ils sont connus sous le nom de chromosomes (Gr-chroma = couleur et Soma = corps). Sutton en 1903, a d'abord suggéré que des gènes ou des facteurs héréditaires sont véhiculés par des chromosomes.
Et en 1871, pour la première fois, isolée par Miescher, son analyse morphologique était beaucoup moins avancée que celle du cytoplasme. Kossel a pu démontrer la présence d'acide nucléique et a reçu le prix Nobel en 1910.
Les cellules sans noyau ont un avenir très limité. Le seul type de cellule animale commun sans noyau, le globule rouge de mammifère, ne vit que 3 à 4 mois; en dehors de son rôle dans le transport de l'oxygène, ses activités métaboliques sont extrêmement limitées.
Les cellules d'œuf dont les noyaux ont été retirés expérimentalement peuvent se diviser pendant un certain temps, mais les produits de la division ne se différencient jamais en types de cellules spécialisées et finissent par mourir. Les fragments sans noyau, coupés dans de grands organismes acellulaires tels que les amibes ou l'algue Acetabularia, survivent temporairement, mais finissent par mourir à moins que des noyaux d'autres cellules ne soient greffés dans ceux-ci.
Le noyau est essentiel à la poursuite à long terme du métabolisme et à la capacité des cellules à modifier de manière significative leur structure et leur fonction (comme dans la différenciation). Cela reflète en grande partie le rôle primordial du noyau dans la production de l'ARN nécessaire à la synthèse des protéines.
Lorsque les cellules changent, de nouvelles fonctions et structures nécessitent de nouvelles protéines. Même les cellules dont le métabolisme et la structure sont constants montrent un remplacement continu (renouvellement) des macromolécules et probablement des organites, y compris des parties de la machinerie synthétisant les protéines du cytoplasme.
Morphologie:
Forme:
En général, le noyau a tendance à être sphérique, mais peut être fusiforme, ellipsoïdal, aplati, en fonction de la forme et de la fonction de la cellule. Dans les cellules jeunes, il est plus souvent sphérique et situé au centre, mais les cellules différenciées peuvent être déplacées et de forme irrégulière.
Le noyau peut être allongé comme dans l'épithélium cylindrique et les cellules musculaires. Dans certains cas, il devient irrégulier et on a souvent observé dans certains cas particuliers que les cellules vivantes, telles que les cellules cartilagineuses, les leucocytes ou les ovules d'animaux, subissent de lents changements de forme amiboïde.
Les noyaux (Noyau singulier) de forme irrégulière ou amoeboide sont fréquents dans les cellules caractérisées par un métabolisme très actif, auquel cas les noyaux sont souvent non seulement de grande taille, mais montrent une augmentation importante de la surface par la formation de lobes, de sacculations, ou même dans les cas extrêmes de ramifications complexes ramifiées à travers la cellule.
Un exemple extrême de cela est fourni par les glandes en rotation de certaines larves d'insectes (Lépidoptères et Trichoptères), dans lesquelles le noyau, à l'origine sphéroïdal, prend finalement une apparence labyrinthique avec des convolutions occupant une grande surface dans la cellule.

Dans certains types de cellules, la surface du noyau peut également être augmentée en se séparant en vésicules plus ou moins séparées ou en caryomérites, formant ainsi un noyau ou un nid nucléaire «polymorphe». Les noyaux de forme vésiculaire ou sphérique sont généralement présents dans les cellules tissulaires de la plupart des animaux et des plantes multicellulaires. Les noyaux massifs se trouvent généralement dans les cellules germinales mâles des animaux en général et de nombreuses plantes inférieures (dans les plantes coénocytaires comme Vaucharia, etc.).
Position:
La position du noyau est déterminée par de nombreuses causes, telles que la tension superficielle, la position de la vacuole, la densité relative du cytoplasme dans différentes parties de la cellule. Dans les cellules embryonnaires, il occupe presque toujours le centre géométrique.
Dans les cellules non vacuolées des plantes, il occupe même le centre de la masse cytoplasmique. Dans les cellules adipeuses, dans les œufs riches en jaune, le noyau est forcé vers la périphérie par l'accumulation du paraplasme. Dans les cellules glandulaires, il est situé en position basale, les granules occupant le cytoplasme apical. La position du noyau est également liée à la fonction de la cellule.
Taille:
En 1895, Boveri a montré que la taille des noyaux chez les larves d'échinoderme dépend du nombre de chromosomes que chacun d'eux contient. Gates 1901, cependant, apporta des preuves montrant que cette règle n'était nullement universelle. La taille du noyau est variable, mais en général elle a une relation avec celle du cytoplasme. Ceci peut être exprimé numériquement dans l'indice dite nucléoplasmique (NP) de O. Hertwigi 1906).
NP = Vn / Vc-Vn
Où Vn est le volume nucléaire et Vc le volume de la cellule.
Cet indice NP montre la relation entre le volume du cytoplasme et celui du noyau, c’est-à-dire que si le premier augmente, le second doit également augmenter. Le rapport entre le volume du noyau et celui du cytoplasme dans une cellule est appelé "rapport nucléoplasmique" ou "rapport caryoplasmique".
Nombre:
Le nombre de noyaux présents dans une masse de protoplasme dépend en grande partie de la masse de la masse puisqu'un certain rapport entre le volume de la surface nucléaire et le volume cytoplasmique doit être maintenu pour que le système tout entier puisse agir correctement.
En général, toutes les cellules sont uninucléées ou mononucléées, c’est-à-dire qu’un noyau est constitué d’une cellule mais que, dans certains cas, l’état binucléé se présente, comme dans Paramecium caudatum. Dans quelques autres cas, la condition poly ou multinucléée existe comme dans Opalina, fibre musculaire striée, etc.
Structure:
Le noyau est composé de quatre composants:
(1) enveloppe nucléaire,
(2) la sève nucléaire,
(3) nucléole, et
(4) Réseau de chromatine.
1. Enveloppe nucléaire:
L'enveloppe nucléaire a récemment fait l'objet de nombreuses enquêtes. Vu avec le microscope à contraste de phase standard, il apparaît comme une ligne plus sombre, cette ligne séparant le contenu nucléaire du cytoplasme. On observe que dans la plupart des cellules, il se décompose et se reforme lorsque la cellule se centre et complète la division. Cependant, chez certaines algues, protozoaires et champignons, il ne se décompose pas pendant la division.
L'observation au microscope optique montre que l'enveloppe nucléaire sert de barrière entre le noyau et le cytoplasme. La manière dont le noyau régule le flux de matière commence tout juste à être étudiée. Des études ultra-structurelles révèlent que l'enveloppe nucléaire est en réalité composée de deux membranes séparées par un espace périnucléaire de 110 à 400 A de large.
La membrane interne semble être en contact avec la chromatine nucléaire, que l'on peut voir se condenser sur sa surface; la membrane externe est continue avec le réticulum endoplasmique cytoplasmique et est souvent observée avec des ribosomes liés à sa surface.
L'association de la membrane externe au RE aboutit à une continuité entre le noyau et le cytoplasme. Cependant, cette continuité n'est pas complète en raison de la présence de la membrane nucléaire interne, qui sert encore de barrière structurelle.
Pores nucléaires :
Étant donné que la synthèse de l'acide ribonucléique a lieu dans le noyau, alors que la synthèse des protéines a lieu dans le cytoplasme, il est évident que les substances doivent pouvoir se déplacer vers l'extérieur. En fait, il existe un flux de matière dans les deux sens à travers les pores nucléaires qui, malgré leur taille, ne sont pas des canaux largement ouverts.
Les deux membranes nucléaires sont fusionnées à intervalles réguliers pour former des pores d'environ 600 ° C de diamètre. Celles-ci ont été vues pour la première fois par Callan et Tomlin (1950). On estime que les pores occupent environ 10% de la surface de la membrane nucléaire ou que le nombre total de pores par noyau peut varier de 100 à 5 x 10 7 . Ces pores apparaissent comme des zones approximativement circulaires ou polygonales.

Cependant, les pores ne sont pas simplement des trous dans la membrane nucléaire. Une grande partie de l'ouverture est occupée par un agencement cylindrique ou annulaire de granules ou de matériau fibrillaire moyennement dense, qui semble être continu avec la lame fibreuse interne.
Le matériau annulaire est organisé comme un complexe de pores. L'anneau est constitué de huit granules annulaires sur sa surface latérale nucléaire et cytoplasmique et d'un granule central au centre. Les fibres s'étendent à partir du granulé central et du matériau annulaire.
Certains matériaux amorphes forment un diaphragme sur le pore. L'importance des diaphragmes et des granules est incertaine car ils ne semblent pas être des composants universels des pores nucléaires. Ce sont peut-être des structures transitoires présentes dans certains états physiologiques et non dans d'autres.
Lamina fibreuse:
Les membranes nucléaires de nombreuses cellules présentent une couche supplémentaire appelée lamina fibreuse. Il s’oppose à l’aspect interne de la membrane nucléaire interne qui fait face à l’intérieur du noyau. Il est composé de fins filaments composés de protéines. Ils fournissent probablement un renforcement mécanique aux membranes nucléaires.
Selon certains travailleurs, la lame fibreuse influence l'échange de matériaux entre le noyau et le cytoplasme. Le degré de développement de la couche fibreuse varie considérablement selon les types de cellules. Dans les cellules de mammifère, il est mince mais très développé chez Amoeba et certains autres invertébrés. En Amoeba, il a une configuration semblable à un nid d'abeilles et a une épaisseur de 1000 à 1500 A °.

Perméabilité de l'enveloppe nucléaire :
Plusieurs expériences suggèrent que les complexes de pores pourraient être des ouvertures temporaires ou permanentes dans l'enveloppe nucléaire. En injectant des particules d'or colloïdale, dont la taille varie de 2, 5 à 17 nm, dans le cytoplasme des amibes, il s'est avéré que celles dont le diamètre atteignait 8, 5 nm pénétraient rapidement dans le noyau.
Les particules de diamètre compris entre 8, 9 et 10, 6 nm ont pénétré plus lentement et les plus grosses n’y sont pas entrées du tout. Ces résultats indiquent que les ouvertures sont inférieures à la taille des pores. Des preuves ont été obtenues avec ces techniques, suggérant que le pore est une voie pour l'échange de macromolécules. Les anneaux peuvent réguler l’échange en fonction de la taille et éventuellement de la nature chimique de la substance pénétrante.
Il est important de considérer que la perméabilité de l'enveloppe nucléaire n'est pas fixe, mais varie selon les types de cellules et au sein d'une cellule donnée, au moins pendant le cycle de division. Ces différences sont imputables à des modifications de la nature du matériau annulaire (Feldher, 1971).
La présence de pores dans l'enveloppe nucléaire doit être corrélée à certaines des propriétés électrochimiques de cette structure, qui peuvent être étudiées à l'aide de microélectrodes linéaires.
Deux types d’enveloppes nucléaires ont été reconnus avec cette technique. Lorsque les cellules géantes de la glande salivaire de Drosophila sont pénétrées avec une microélectrode, il se produit un changement de potentiel brusque au niveau de la membrane plasmique (-12 mV); puis, lorsque la microélectrode entre dans le noyau, il se produit une autre baisse de potentiel négatif au niveau de la membrane nucléaire (-13 mV).
Ces résultats suggèrent que l'enveloppe nucléaire peut constituer une barrière de diffusion pour des ions aussi petits que K +, Na + ou Cl. Cependant, dans l'enveloppe nucléaire présente dans les occultes, il n'y a pas de potentiel détectable, ce qui indique un échange libre d'ions entre le noyau et le cytoplasme.

Le mécanisme par lequel le passage de ces substances est obtenu est inconnu. Des études cytochimiques ont toutefois révélé la présence d’une ATPase dans les pores pouvant fournir l’énergie nécessaire au transfert de macromolécules.
Origine de la membrane nucléaire :
La membrane nucléaire est considérée comme une structure cytoplasmique spécialisée issue du système membranaire cytoplasmique. Au cours de la télophase, il se forme par l'accumulation de vésicules de réticulum endoplasmique autour du groupe de chromosomes, qui ensuite fusionnent pour former une membrane complète.
Il existe essentiellement deux théories sur la manière dont l’enveloppe nucléaire est reconstituée après la télophase: la paire de lamelles est synthétisée de novo sur les sites où chromosomes et cytoplasme sont en contact (Jones 1960) ou des parties du réticulum endoplasmique déjà présent sont adjacentes. le noyau de la télophase et fusionner pour former l'enveloppe nucléaire.
Dans les spermatocytes de criquets, par exemple, les vésicules de 0, 5 ц qui se forment à la surface des chromosomes ne peuvent être distinguées des vésicules du réticulum endoplasmique (Bahr, 1959, p. 61). Ils se rangent progressivement de manière à être parallèles à la surface du noyau, puis fusionnent pour former une enveloppe nucléaire continue.
2. SAP nucléaire:
Une masse acidophile légère ou peu colorée, le «caryolymphe» ou «sève nucléaire», remplit complètement l’espace nucléaire où se trouvent les autres composants. C'est Claude qui en 1943 a précipité la sève nucléaire par un fixateur histologique et coloré avec des colorants acides. Dans les œufs et dans les grandes cellules (p. Ex. Acetabularia), la sève nucléaire est clairement visible. Selon Stick (1951, 58), il donne un test cytochimique positif pour l'ARN, les groupes de protéines, y compris SH et les glycoprotéines.
De nombreuses voies métaboliques ont également été mises en évidence dans la sève nucléaire, semblable au cytoplasme. Ceux-ci incluent la glycolyse, le shunt hexose monophosphate, le cycle de l'acide citrique, etc. Le shunt hexose monophosphate est important car il alimente le noyau en pentose. En outre, dans le noyau est synthétisé le NAD + qui est le coenzyme de nombreuses déshydrogénases. Les enzymes caractéristiques du noyau sont l'ADN et les ARN-polymérases.
Composition chimique du noyau:
Les composants chimiques suivants ont été analysés à partir de la matière nucléaire isolée:
(i) Nucléoprotéines
(a) Nucléoprotamine
(b) Nucléo-histones
(c) Protéines non-histones ou acides.
(ii) acides nucléiques.
(iii) Enzymes.
(iv) les lipides.
(i) Nuceoproteins:
Outre les acides nucléiques, l’autre constituant important du noyau de la cellule est constitué par les composants protéiques. Les chromosomes des plantes et des cellules animales, contrairement au matériel génétique des bactéries, consistent en ADN associé à des protéines. Dans des chromosomes polygéniques géants, des protéines ont été trouvées dans la bande et dans les régions intégrandes de l’activité des gènes.
La protéine est associée aux boucles prolongées des chromosomes du lampbrush. Les nucléoles contiennent de fortes proportions de protéines et il y a bien sûr des protéines présentes dans la sève ou le nucléoplasme nucléaire et, en association avec les lipides, dans les membranes nucléaires.
La partie protéique du noyau est certainement très complexe et comporte plusieurs composants. Parmi celles-ci, les plus connues sont deux protéines très basiques et simples: les protamines et les histoires. En plus de celles-ci, il existe des protéines acides, des protéines dites non histones et des enzymes.
a) Protamine ou nucléoprotamine:
C'est Miescher qui a découvert la protamine dans le sperme de saumon. Ce sont des protéines basiques simples ayant un très faible poids moléculaire. Celles-ci sont très riches en arginine, un acide aminé basique, que l'on retrouve dans les spermatozoïdes de certains poissons et qui sont étroitement liées à l'ADN par le biais d'une liaison de sel.
Comme les protamines sont plus petites que les histones, cela signifie que les chromosomes et les noyaux peuvent être entassés dans un espace plus petit, permettant une plus grande mobilité du sperme. En général, ils consistent en environ 28 polypeptides d'arginine, d'une longueur totale de 100 ° et contiennent 19 arginines et 8 ou 9 acides aminés non basiques. Au cours du développement, il y a remplacement progressif des histones par la protamine. Cela peut être dû à la plus grande affinité de la protamine pour l’ADN.
b) Nucleo-hostones :
Des histones ont été découverts dans les érythrocytes d'oie par Kossel et, peu de temps après, dans le thymus par Lilientfeld. Des histones ont également été trouvés dans certains noyaux de cellules végétales, notamment des embryons de blé et de noix de cèdre de Mirsky.
Les nucléoprotéines sont également des protéines basiques ayant un poids moléculaire d'environ 12 000. En plus d'environ 13% d'arginine, les histones contiennent d'autres résidus basiques, notamment la lysine et l'histidine. Plusieurs histones de composition différente ont été isolées et trois types ont été caractérisés.
(i) très riche en lysine.
(ii) riche en arginine.
(iii) et légèrement riche en lysine.
Ceux-ci sont hétérogènes et se composent de plusieurs composants. Les histones se trouvent dans tous les noyaux des organismes supérieurs, bien qu'ils aient été peu étudiés chez les plantes. Celles trouvées dans les plantes ressemblent toutefois aux histones trouvées chez les vertébrés.
La principale fonction de l'histone attribuée est d'agir comme une «colle» chromosomique liant ensemble les unités génétiques de l'ADN. Il est également connu que l'ADN seul et que les protéines (principalement les histones) du complexe nucléo-protéique peuvent protéger partiellement l'ADN des dommages causés par les radiations.
Les preuves indiquent maintenant, cependant, que le rôle vraiment important joué par les histones réside dans la représentation de l'activité génétique des cellules. En 1950, Stedman et Stednian suggèrent que les histones interagissent de manière spécifique avec l'ADN, l'empêchant ainsi d'agir comme un tamplate pour la synthèse d'ARN et empêchant ainsi le transfert d'informations génétiques au cytoplasme.
(c) Non-histones ou protéines acides :
L'analyse chimique de fils de nuceli et de chromatine isolés a permis de mettre en évidence un autre type de protéine, généralement appelé protéine non histone. Cette protéine contient du tryptophane et possède des propriétés acides.
Une autre fraction non histone a été identifiée à partir de fils de chromatine isolés. Le résidu insoluble apparaît au microscope sous forme de fils en spirale qui conservent les caractéristiques des chromosomes en interphase. D'un point de vue chimique, il est très intéressant que ces chromosomes résiduels contiennent plus d'acide ribonucléique (ARN) que d'ADN.
La quantité considérable de protéines non histones présentes dans une cellule métaboliquement active contraste nettement avec la composition du spermatozoïde, qui est beaucoup moins active.
ii) acides nucléiques:
Les acides nucléiques, qui sont de deux types: l'acide ribonucléique (ARN) et l'acide désoxyribonucléique (ADN) ont été observés dans la région nucléaire de la cellule.
(iii) Enzymes nucléaires:
Un certain nombre d'enzymes ont été trouvées dans les vésicules germinales isolées d'ovocytes de grenouille par Brachet et Dupsiva. Leur concentration réelle est difficile à déterminer en raison de la grande perméabilité de la membrane nucléaire et les enzymes risquent également de fuir des vésicules germinales isolées lors de leur isolement.
Selon des études récentes, les enzymes nucléaires appartiennent à deux classes. Certains ont une distribution générale alors que d'autres se trouvent préférentiellement dans certains tissus. Dans le premier groupe, seules certaines enzymes liées au métabolisme des nucléosides, telles que l'adénosine diaminase, la nucléoside phosphorylase et la guanase se trouvent en forte concentration.
D'autres enzymes comme les estérases sont présentes à des concentrations variables. D'autres encore, telles que la phosphatase alcaline, les nucléotide phosphatases et la β-glucoronidase sont soit présentes à faible concentration, soit totalement absentes. Parmi les enzymes spéciales, la catalase et l'arginase semblent être concentrées dans certains noyaux mais font défaut dans d'autres.
iv) Lipides nucléaires:
La teneur en lipides du noyau a été étudiée dans des noyaux isolés. Récemment, il a été rapporté que le complexe lipo-protéique dans les noyaux d'érythrocytes de rate et de poulet était à environ 10% de lipides. Ceux-ci donnent des tests positifs pour les phospholipides et le cholestérol.
3. Nucléole:
Le noyau contient un gros granule dense, sphérique et acidophile, appelé nucléole. Les nucléoles ont été découverts par Fontona en 1781. Il était bien établi au 19ème siècle que la taille du nucléole est liée à l'activité synthétique de la cellule.
Par conséquent, les cellules avec peu ou pas d'activités de synthèse, par exemple les spermatozoïdes, les blastomères, les cellules musculaires, etc., contiennent des nucléoles plus petits ou nuls, alors que les ovocytes, les neurones et les cellules sécrétoires qui synthétisent les protéines ou d'autres substances contiennent comparativement nucléoles de grande taille.
Le nombre de nucléoles dans le noyau dépend de l'espèce et du nombre de chromosomes. Le nombre de nucléoles dans le noyau des cellules peut être un, deux ou quatre. La position du nucléole dans le noyau est excentrique.
Structure fine du nucléole :
En ce qui concerne la structure fine des nucléoles, Estable et Sotelo (1955) ont rapporté qu'ils sont formés de deux parties: un nucléolonème filamenteux et un pars amorpha. On dit que le nucléolonème se divise et se répartit de manière égale dans la mitose fille et il est considéré comme une structure permanente qui persiste pendant toute la mitose en association avec le chromosome.

On pense que la partie filamenteuse des nucléoles contient de l'ADN, tandis que le pars amorpha est partiellement constitué d'ARN. Pars amorpha subit le cycle caractéristique de formation à la télophase et de disparition à la prophase.
L’introduction de nouveaux milieux et techniques d’inclusion a permis une meilleure analyse de l’organisation nucléolaire.
Ainsi, quatre composantes principales peuvent être reconnues:
1. Une portion fibrillaire d'environ 50A ° de diamètre et jusqu'à 300-400 A ° de long (Marrinozzi, 1963). Dans certains cas, on peut constater que ces fibriles ont une structure à double brin (Terzakis, 1965). Ces structures ont également été décrites comme des éléments tubulaires de 15 A 0 de diamètre.
2. Une portion granulaire composée de granules denses d'un diamètre moyen de 150 à 200 °, plus ou moins nombreux le long des brins du réseau fibrillaire et entre ceux-ci (Marrinozzi, 1963).
3. Une région amorphe de faible électrondensité, présente dans certains noyaux (Terzakis, 1965) en protéine.
4. La chromatine nucléolaire associée située autour du nucléole et ayant fréquemment des composants intra-nucléolaires semble être constante dans les cellules de vertébrés, mais leurs quantités respectives peuvent varier.
Chimie :
Des études cytochimiques indiquent que 5 à 10% du nucléole est constitué d’ARN; le reste étant la protéine. Les principales composantes protéiques sont les phospho-protéines. Aucun son n'a été trouvé dans les nucléoles isolées.
Il existe des preuves suggérant la présence d'orthophosphate, qui pourrait servir de précurseur de l'ARN phosphore. Le contenu en enzymes du nucléole est peu connu. Cependant, la présence de phosphatase acide, de nucléoside phosphorylase et d’enzymes synthétisant NAD + a été mise en évidence. L'ARN méthylase a également été localisée dans le nucléole de certaines cellules.
L'ADN est absent. Le nucléole peut être entouré d'un anneau de chromatine Feulgen-positive qui représente en réalité les régions hétérochromatiques des chromosomes.
Biogenèse du cycle nucléole ou nucléolaire:
Le nucléole en tant que corps organisé manque de continuité. Il disparaît au début de la division cellulaire (prophase) et réapparaît à la fin de la division cellulaire au stade télophase.
Un nucléole est formé dans une région définie par un ou plusieurs chromosomes d'un ensemble de chromosomes haploïdes. Ces chromosomes sont connus sous le nom de chromosomes nucléolaires.
La majorité des espèces diploïdes possèdent deux chromosomes nucléolaires dans chaque cellule diploïde ou somatique. Mais chez l'homme, les chromosomes numérotés 13, 14, 15, 21 et 22 participent à la formation du nucléole.
La région spécifique de ces chromosomes actifs dans la formation nucléolaire est appelée zone nucléolaire organisatrice nucléolaire. Très souvent mais pas toujours, il est marqué par une constriction secondaire. L'organisateur nucléolaire porte les gènes de l'ARN ribosomal 18S et 28S.
La constriction secondaire avec l'organisateur nucléolaire est morphologiquement différente des autres constrictions secondaires. Au stade pachytène de Zea mays, un corps organisateur nucléolaire sombre est associé à la zone nucléolaire.
Types de nucléoles :
Sur la base de la distribution des granules et des fibrilles de ribonucléoprotéines, on peut distinguer trois types de nucléoles (Wilson).
1. Nuclcoli avec nucléolonème, que l'on trouve dans la plupart des cellules appelées plasmosomes.
2. Des nucléoles compactes sans nucléolonème, décrites dans la glande salivaire des sciuridés et dans le protozoaire, Tetrahymena pyriformis. Dans ces nucléoles, les granules et les fibrilles de ribonucléoprotéine sont uniformément répartis.
3. Des nucléoles en forme d'anneau avec des granules et des fibrilles de ribounoprotéine, présents uniquement dans la région périphérique (nudeolonema périphérique), ont été rapportés dans des cellules endothéliales, des cellules musculaires lisses et des cellules lymphosarceuses. Dans ces nucléoles, la région centrale est en grande partie constituée de chromatine.
Les fonctions:
(i) production d'ARN :
Le nucléole est l’un des sites les plus actifs de la synthèse d’ARN. Il produit environ 70 à 90% d'ARN cellulaire dans de nombreuses cellules. C'est la source de l'ARN ribosomal (ARNr). La chromatine du nucléole contient des gènes ou l'ADN ribosomal (ADNr) permettant de coder l'ARN ribosomal.
Les fibrilles représentent l'origine de l'ARN ribosomal et les granules l'étape suivante. Les granules sont à leur tour les précurseurs des ribosomes. Le nucléole fabrique ainsi des précurseurs du ribosome plutôt que des ribosomes entiers.
Chromation → Fibrilles → Granulés → Ribosomes
(Contenant de l'ADN) (contenant de l'ARN) (contenant de l'ARN).
Le nucléole peut également produire certains types d’ARN messager (ARNm) et au moins un type d’ARN de Wight de faible poids moléculaire.
(ii) synthèse des protéines :
Maggio (1960) et d'autres ont suggéré que la synthèse des protéines avait lieu dans le nucléole. Si cela est vrai, les protéines ribosomales sont produites dans le nucléole. D'autres études suggèrent cependant que les protéines ribosomales sont synthétisées dans le cytoplasme.
(iii) Formation de ribosome :
Chez les eucaryotes, le gène codant pour l'ARN contient une chaîne d'au moins 100 à 1 000 copies répétées de l'ADN. Cet ADN se dégage sous forme de boucles issues de la fibre chromosomique. Les boucles d’ADN sont associées à des protéines pour former des nucléoles.
L'ADN sert de matrice pour l'ARNr 45S. La moitié de l'ARNr 45S est décomposé en ARNr 28S et 18S. L'autre moitié est décomposée au niveau des nucléotides. À l'intérieur du nucléole, l'ARNr 28S se combine avec des protéines produites dans le cytoplasme pour former la sous-unité ribosomale 60S. L'ARNr 18S s'associe également à des protéines pour former la sous-unité 40S du ribosome.
4. Réseau de chromatine :
La chromatine se présente sous la forme d'une structure filiforme, enroulée et allongée Celles-ci sont colorées avec des colorants basiques tels que la coloration de Feulgen, l'acétocarmine, etc.
Celles-ci sont visibles pendant la phase d'interphase. Au cours de la division cellulaire, ils deviennent des structures épaisses en forme de ruban appelées chromosomes.
Signification du noyau:
Expérience de Hammerling:
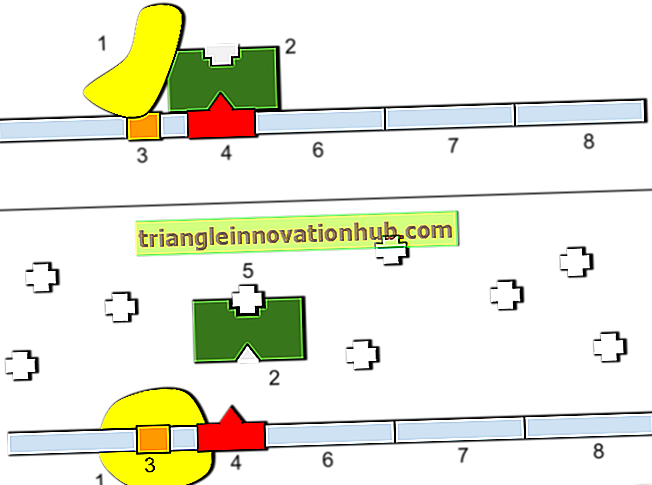
J. Hammerling, un biologiste allemand, a prouvé de manière expérimentale la signification du noyau. Si un fragment contenant le noyau est coupé d'Acetabularia d'une espèce, caractérisé par une morphologie donnée, le fragment régénérera une cellule entière de cette espèce.
Cette capacité de régénération permet des expériences du type illustré à la figure 8.6 dans lesquelles des noyaux d'une espèce sont combinés à un cytoplasme d'espèces différentes. Hammerling a mené certaines expériences en utilisant deux espèces d'une algue verte, Acetabularia. Les deux espèces, à savoir A. crénulées et A, méditerranéenne utilisées dans cette expérience se différencient par la forme de leurs calottes. Chez A. crenulata, le chapeau a des rayons lâches, alors que chez A. mediterranea, on trouve un chapeau ressemblant à un parapluie.

Le noyau dans les deux espèces est situé dans le rhizoïde au bas de la tige. Si le capuchon est coupé, il se développera à nouveau et sa forme sera celle du type original. Cependant, si après le retrait des capuchons, une tige d'une espèce est greffée sur du rhizoïde (contenant le noyau) de l'autre espèce, la forme de la coiffe sera déterminée par le noyau et non par la tige. Si le noyau appartient à A. crenulata, la forme de la coiffe sera du type crenulata. Si le noyau provient de A. mediterranea, la coiffe sera de type mediterranea.
Lorsque les deux noyaux sont présents, la forme de la coiffe sera intermédiaire. La conclusion tirée de telles expériences est que le noyau produit un matériau qui pénètre dans le cytoplasme et participe au contrôle de la croissance et de la morphologie cellulaires.
La découverte cruciale est que la morphologie des cellules régénérées devient pratiquement identique à celle de l'espèce dont le noyau est extrait. Dans les fragments hybrides avec le noyau d’une espèce et la majeure partie du cytoplasme de l’autre espèce, l’ancien matériel cytoplasmique persiste pendant un certain temps et peut influencer la forme des cellules. Cependant, il est finalement épuisé et remplacé par du matériel nouvellement produit à partir du noyau.